Soortvorming
Speciatie gaat over hoe soorten zich vormen. Het is een belangrijk onderdeel van de evolutiebiologie. Darwin dacht dat de meeste soorten rechtstreeks voortkwamen uit reeds bestaande soorten. Dit wordt anagenese genoemd: soorten door verandering,…
Speciatie gaat over hoe soorten zich vormen. Het is een belangrijk onderdeel van de evolutiebiologie.
Darwin dacht dat de meeste soorten rechtstreeks voortkwamen uit reeds bestaande soorten. Dit wordt anagenese genoemd: soorten door verandering, of "fyletische evolutie". Gedurende een groot deel van de 20e eeuw dachten wetenschappers dat de meeste soorten ontstonden door afsplitsing van eerdere soorten. Dit wordt cladogenese genoemd. De algemene opvatting was dat de meeste splitsingen van soorten veroorzaakt of op weg geholpen worden door isolerende mechanismen.
Ongetwijfeld is de fysieke scheiding van soorten die ooit samenleefden een belangrijke factor. Dit wordt geïllustreerd door vele voorbeelden, waarvan er hieronder enkele worden besproken.
Werk in de afgelopen 20 jaar heeft echter een aantal andere oorzaken aangetoond. Uit analyse van de DNA-sequentie van levende wezens is gebleken dat er vaak sprake is van enige hybridisatie tussen verwante soorten. Dat betekent dat door deze kruisingen genen zijn overgedragen. Dat betekent dan weer dat reproductieve isolatie niet de enige definitie van een soort is, en dat voor soortvorming niet altijd allopatrie nodig is (soorten die reproductief gescheiden zijn). De onderstaande paragrafen illustreren het idee dat fysieke scheiding van primair belang was bij de vorming van nieuwe soorten.
Afbeeldingengalerij
10 Afbeeldingen









Isolerende mechanismen
Isolerende mechanismen zijn zaken die een succesvolle voortplanting tussen groepen in een soort verhinderen. Reproductieve isolatie van populaties komt tot stand. Dit is bijzonder belangrijk voor het biologische soortenconcept, aangezien soorten worden gedefinieerd door reproductieve isolatie.
Isolerende mechanismen kunnen worden onderverdeeld in twee groepen, vóór en na de bevruchting.
Vóór de bevruchting
Factoren die verhinderen dat individuen paren.
- Geografische isolatie: Soorten komen voor in verschillende gebieden, en zijn vaak gescheiden door barrières.
- Temporele isolatie: Individuen paren niet omdat ze op verschillende tijden actief zijn. Dit kunnen verschillende momenten van de dag of verschillende seizoenen zijn. De paringsperioden van de soorten komen niet altijd overeen. Individuen ontmoeten elkaar niet tijdens hun paringsperiode, of helemaal niet.
- Ecologische isolatie: Individuen paren alleen in hun favoriete habitat. Ze komen geen individuen tegen van andere soorten met andere ecologische voorkeuren.
- Gedragsisolatie: Individuen van verschillende soorten kunnen elkaar ontmoeten, maar men herkent eventuele seksuele signalen niet. Een individu kiest in de meeste gevallen een lid van zijn eigen soort.
- Mechanische isolatie: Copulatie kan worden geprobeerd, maar de overdracht van sperma vindt niet plaats. De individuen kunnen onverenigbaar zijn vanwege grootte of morfologie.
- Gametische incompatibiliteit: Er vindt een zaadoverdracht plaats, maar de eicel wordt niet bevrucht.
Na de bevruchting
Factoren die een succesvolle paring verhinderen, zoals genetische incompatibiliteit, hybriditeit of steriliteit.
- Zygotische sterfte: De eicel wordt bevrucht, maar de zygote ontwikkelt zich niet.
- Hybride invabiliteit: Hybride embryo wordt gevormd, maar is niet levensvatbaar.
- Hybride steriliteit: De hybride is levensvatbaar, maar het resulterende volwassen dier is steriel.
- Afbraak van de hybride: Hybriden van de eerste generatie (F1) zijn levensvatbaar en vruchtbaar, maar verdere hybride generaties (F2 en terugkruisingen) zijn levensvatbaar of steriel.

Geografische isolatie
Dit wordt beschouwd als de meest voorkomende oorzaak van soortvorming. De eerste die hieraan dacht was Moritz Wagner, een Duitse ontdekkingsreiziger en natuurhistoricus.
Wagners vroege carrière was als geograaf, en hij publiceerde een aantal geografische boeken over Noord-Afrika, het Midden-Oosten en Tropisch Amerika. Hij was ook een enthousiast naturalist en verzamelaar, en onder biologen is hij vooral bekend om dit werk. Ernst Mayr, de evolutionist en historicus van de biologie, heeft de betekenis van Wagner beschreven.p562–565
Gedurende zijn drie jaar in Algerije bestudeerde Wagner (onder andere) de loopkevers Pimelia en Melasoma. Elk geslacht is verdeeld in een aantal soorten, die elk beperkt zijn tot een stuk van de noordkust tussen de rivieren die van het Atlasgebergte afdalen naar de Middellandse Zee. Zodra men een rivier oversteekt, verschijnt een andere, maar nauw verwante soort.
"... een beginnende soort zal alleen [ontstaan] wanneer een paar individuen de grenzen van hun verspreidingsgebied overschrijden... de vorming van een nieuw ras zal nooit slagen... zonder een lange voortdurende scheiding van de kolonisten van de andere leden van hun soort".
Dit was een vroege beschrijving van een proces van één soort geografische speciatie. In 1942 werd het opnieuw geïntroduceerd door Mayr, en het belang van geografische speciatie werd een van de kernideeën van de evolutionaire synthese.
Een andere term voor geografische speciatie is allopatrische speciatie. Allopatrie betekent "verschillend land".
Vrije eilanden
Vulkanische eilanden worden gevormd zonder leven, en al het leven moet door wind of water worden aangevoerd. Van de Hawaiiaanse eilanden en de Galapagoseilanden weten we dat alle levensvormen veranderen wanneer ze de eilanden vanaf het vasteland bereiken.
2Op ongeveer 17.000 km2 hebben de Hawaïaanse eilanden de meest diverse verzameling drosophiliden ter wereld, die leven van regenwouden tot bergweiden. Er zijn ongeveer 800 soorten Hawaiiaanse drosophiliden bekend.
Studies tonen een duidelijke "stroom" van soorten van oudere naar nieuwere eilanden. Er zijn ook gevallen van kolonisatie terug naar oudere eilanden, en het overslaan van eilanden, maar die komen veel minder vaak voor.
Volgens radioactieve datering met kalium/argon dateren de huidige eilanden van 0,4 miljoen jaar geleden (mya) (Mauna Kea) tot 10 mya (Necker). Het oudste lid van de Hawaïaanse archipel dat nog boven zee ligt, is Kure Atoll, dat kan worden gedateerd op 30 mya.
De archipel zelf, ontstaan doordat de Pacifische plaat over een hete plek bewoog, bestaat al veel langer, ten minste tot in het Krijt. De Hawaiiaanse eilanden plus voormalige eilanden die nu onder de zee liggen, vormen de keten van de Hawaiian-Emperor seamounts; en veel van de onderwaterbergen zijn guyots.
Alle inheemse drosophilide soorten in Hawaiʻi stammen blijkbaar af van één enkele voorouderlijke soort die de eilanden ongeveer 20 miljoen jaar geleden koloniseerde. De daaropvolgende adaptieve radiatie werd gestimuleerd door een gebrek aan concurrentie en een grote verscheidenheid aan lege niches. Hoewel het mogelijk is dat één enkel zwanger vrouwtje een eiland koloniseert, is het waarschijnlijker dat het om een groep van dezelfde soort gaat.
Er zijn andere dieren en planten op de Hawaïaanse archipel die soortgelijke, zij het minder spectaculaire, aanpassingsstralingen hebben ondergaan.
Ringsoorten
In de biologie is een ringsoort een verbonden reeks naburige populaties, die zich elk kunnen kruisen met naburige populaties. De twee uiteinden van de keten overlappen elkaar.
De twee eindpopulaties in de reeks zijn te ver verwant om met elkaar te kruisen. Dergelijke niet-voortplantende, maar genetisch verbonden "eindpopulaties" kunnen in hetzelfde gebied naast elkaar voorkomen en zo een "ring" vormen.
Ringsoorten vormen een belangrijk bewijs van evolutie: zij illustreren wat er in de loop der tijd gebeurt als populaties genetisch van elkaar verschillen. Richard Dawkins merkte op dat ringsoorten "ons alleen in de ruimtelijke dimensie iets laten zien wat altijd in de tijdsdimensie moet gebeuren".
Het is echter moeilijk om een eenvoudig, rechtlijnig voorbeeld te vinden.
Larus meeuwen
Dit was het klassieke voorbeeld van ringsoorten. Het verspreidingsgebied van deze meeuwen vormt bijna een ring rond de Noordpool (die normaal gesproken niet door meeuwen wordt overvlogen). De kleine mantelmeeuwen en zilvermeeuwen zijn zo verschillend dat ze normaal gesproken niet hybridiseren; er werd dus gezegd dat de groep meeuwen een continuüm vormt, behalve waar de twee lijnen elkaar in Europa ontmoeten. Men is het er echter over eens dat dit niet helemaal juist is, hoewel de details uiterst gecompliceerd zijn.
Ensatina salamanders
De Ensatina-salamander is een ringsoort in de bergen rond de Californische Centrale Vallei. Het complex vormt een hoefijzervorm rond de bergen. Hoewel kruising tussen elk van de 19 populaties rond het hoefijzer mogelijk is, kan de ondersoort Ensatina eschscholtzii aan het westelijke uiteinde van het hoefijzer zich niet vermengen met de Ensatina klauberi aan het oostelijke uiteinde. Het is een illustratie van "bijna alle stadia in een speciatieproces" (Dobzhansky). Richard Highton betoogde dat Ensatina een geval is van meerdere soorten en niet van een continuüm van één soort.
De groenling
De groenling (Phylloscopus trochiloides) kent een aantal ondersoorten, waarvan P. t . viridianus de meest bekende is in Europa. Het is een ringsoort met populaties die ten oosten en westen van het Tibetaanse Plateau uiteenlopen en later aan de noordkant samenkomen. Hun relaties zijn tamelijk verwarrend.




Sympatrische soortvorming
Sympatrische soortvorming verwijst naar de vorming van twee of meer afstammende soorten van één voorouderlijke soort die allemaal dezelfde geografische locatie innemen. Dit wordt nu als zeer gebruikelijk beschouwd.
Bij sympatrische soortvorming divergeren soorten terwijl ze op dezelfde plaats leven. Vaak genoemde voorbeelden van sympatrische soortvorming zijn insecten die afhankelijk worden van verschillende waardplanten in hetzelfde gebied.
Het bestaan van sympatrische soortvorming als speciatiemechanisme werd fel betwist. Men betoogde dat de bewijzen van sympatrische soortvorming in feite voorbeelden zijn van microgeografische soortvorming. In het algemeen is dit nu niet de voorkeursverklaring. Het is veeleer het resultaat van hybridisatie tussen nauw verwante soorten, gevolgd door natuurlijke selectie op de nakomelingen van dergelijke kruisingen. Een algemeen aanvaard voorbeeld van sympatrische soortvorming is dat van de cichliden van het Nabugabo-meer in Oost-Afrika, dat vermoedelijk het gevolg is van seksuele selectie.
Speciatie via polyploïdisatie
Polyploïdie heeft veel snelle soortvorming veroorzaakt, omdat nakomelingen van bijvoorbeeld tetraploïde x diploïde paringen vaak resulteren in triploïde steriele nakomelingen.
Niet alle polyploïden zijn echter reproductief geïsoleerd van hun ouderplanten, en er kan nog steeds genenstroom optreden, bijvoorbeeld door triploïde hybride x diploïde paringen die tetraploïden voortbrengen.
Veel van de bestaande plantensoorten en de meeste diersoorten hebben in hun evolutionaire geschiedenis kennelijk polyploïdisatie ondergaan. De voortplanting van succesvolle polyploïde soorten is soms ongeslachtelijk, door parthenogenese. Om onbekende redenen zijn veel aseksuele organismen polyploïde.
Meidoornvlieg
Een voorbeeld van evolutie aan het werk is het geval van de meidoornvlieg, Rhagoletis pomonella, die sympatrische soortvorming lijkt te ondergaan.
Verschillende populaties van de meidoornvlieg voeden zich met verschillende vruchten. In Noord-Amerika ontstond een aparte populatie in de 19e eeuw, enige tijd nadat appels, een niet-inheemse soort, werden geïntroduceerd. Deze populatie voedt zich normaal gesproken alleen met appels en niet met de historische voorkeur voor meidoorns. De huidige populatie die zich met meidoorns voedt, voedt zich normaal gesproken niet met appels.
Er zijn aanwijzingen dat er sympatrische soortvorming plaatsvindt. Meidoornvliegen worden later in het seizoen volwassen en doen er langer over dan appelvliegen; en er is weinig bewijs van kruising (onderzoekers hebben een hybridisatiegraad van 4-6% gedocumenteerd).
De opkomst van de nieuwe meidoornvlieg is een voorbeeld van evolutie in uitvoering.
Hybridisatie
In zeldzame gevallen ontstaat een nieuwe soort wanneer individuele leden van verschillende soorten met elkaar paren. Meestal zijn de producten van dergelijke hybride paringen onvruchtbaar (niet vruchtbaar), of relatief onvruchtbaar, en worden dus geëlimineerd door natuurlijke selectie.
Er is een voorbeeld gevonden van een nieuwe en succesvolle hybride soort. De nieuwe soort is een hybride van de Italiaanse en Spaanse mussen, en de basisgegevens zijn gecontroleerd door sequentieanalyse van het DNA in hun bloed. De vogel leeft in Italië in een regio waar beide oudersoorten voorkomen. Hij plant zich niet voort met de Spaanse mus, ook al leeft hij met hen samen.
Kunstmatige speciatie
Er zijn nieuwe soorten ontstaan door het houden van gedomesticeerde dieren, maar de oorspronkelijke data en methoden zijn niet duidelijk. Tamme schapen bijvoorbeeld werden gecreëerd door hybridisatie, en produceren geen levensvatbare nakomelingen meer met Ovis orientalis, een soort waarvan zij afstammen.
Tamme runderen daarentegen kunnen worden beschouwd als dezelfde soort als verschillende soorten wilde ossen, gaur, yak, enz. omdat zij daarmee gemakkelijk vruchtbare nakomelingen produceren.
Labsoorten
De best gedocumenteerde creaties van nieuwe soorten in het laboratorium werden eind jaren tachtig uitgevoerd. William Rice en G.W. Salt kweekten fruitvliegjes, Drosophila melanogaster, met behulp van een doolhof met drie verschillende habitatkeuzes, zoals licht/donker en nat/droog.
Elke generatie werd in het doolhof geplaatst, en de groepen vliegen die uit twee van de acht uitgangen kwamen werden apart gezet om zich in hun respectieve groepen met elkaar voort te planten. Na vijfendertig generaties waren de twee groepen en hun nakomelingen reproductief geïsoleerd vanwege hun sterke habitatvoorkeur: ze paren alleen binnen de gebieden die hun voorkeur genieten, en dus niet met vliegen die de voorkeur geven aan de andere gebieden. De geschiedenis van dergelijke pogingen wordt beschreven in Rice en Hostert (1993).
Diane Dodd kon ook aantonen hoe voortplantingsisolatie zich kan ontwikkelen uit paringsvoorkeuren bij Drosophila pseudoobscura na slechts acht generaties met behulp van verschillende soorten voedsel, zetmeel en maltose.
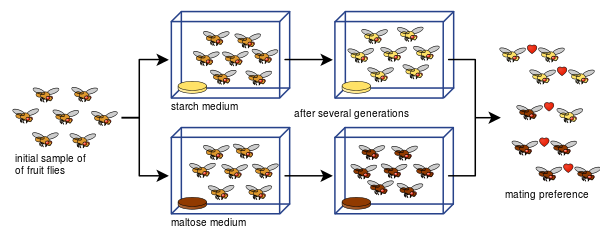
Het experiment van Dodd is door vele anderen gemakkelijk te repliceren, ook met andere soorten fruitvliegen en voedsel.
Versterking (Wallace-effect)
Versterking is het proces waarbij natuurlijke selectie de reproductieve isolatie vergroot.
Het kan voorkomen nadat twee populaties van dezelfde soort van elkaar zijn gescheiden en vervolgens weer met elkaar in contact komen. Als hun voortplantingsisolatie volledig was, hebben ze zich al ontwikkeld tot twee afzonderlijke, onverenigbare soorten.
Als hun reproductieve isolatie onvolledig is, zullen verdere paringen tussen de populaties hybriden voortbrengen, die al dan niet vruchtbaar kunnen zijn. Als de hybriden onvruchtbaar zijn, of vruchtbaar maar minder fit dan hun voorouders, dan is er geen verdere reproductieve isolatie en heeft er in wezen speciatie plaatsgevonden (bijv. zoals bij paarden en ezels).
De redenering hierachter is dat als de ouders van de hybride nakomelingen elk natuurlijk geselecteerde eigenschappen hebben voor hun eigen verschillende omgevingen, de hybride nakomelingen eigenschappen van beide zullen dragen, en niet zo goed in één van beide niches zouden passen als beide ouders. Door de lage fitness van de hybriden zou selectie assortatieve paring in de hand werken, waardoor hybridisatie zou afnemen.
Dit wordt soms het Wallace-effect genoemd, naar de evolutiebioloog Alfred Russel Wallace die eind 19e eeuw suggereerde dat dit een belangrijke factor zou kunnen zijn bij soortvorming.
Als de hybride nakomelingen fitter zijn dan hun voorouders, zullen de populaties weer samensmelten tot dezelfde soort binnen het gebied waar ze met elkaar in contact komen.
Versterking is nodig voor scheiding wanneer er een "hybride zone" is tussen twee vormen van een soort. Hybride zones zijn gebieden waar uiteenlopende populaties elkaar ontmoeten en kruisen. Hybride nakomelingen komen vaak voor in deze gebieden, die meestal ontstaan doordat uiteenlopende soorten met elkaar in contact komen.
Zonder versterking zouden de twee populaties of soorten oncontroleerbare kruisingen hebben. Versterking kan worden geïnduceerd in kunstmatige selectie-experimenten zoals hierboven beschreven.
Vragen en antwoorden
V: Wat is soortvorming?
A: Speciatie is het proces van de vorming van soorten. Het is een belangrijk onderdeel van de evolutiebiologie.
V: Wat dacht Darwin over de vorming van soorten?
A: Darwin dacht dat de meeste soorten rechtstreeks voortkwamen uit reeds bestaande soorten, wat hij anagenese of "fyletische evolutie" noemde.
V: Hoe werd in de 20e eeuw over het algemeen tegen soortvorming aangekeken?
A: In de 20e eeuw dachten wetenschappers dat de meeste soorten werden gevormd wanneer eerdere soorten zich splitsten, wat bekend stond als cladogenese. Men geloofde ook dat deze splitsing werd veroorzaakt of geholpen door isolerende mechanismen zoals fysieke scheiding.
V: Hoe heeft recent onderzoek ons begrip van soortvorming veranderd?
A: Recent onderzoek heeft aangetoond dat hybridisatie tussen verwante soorten kan plaatsvinden en dat via deze kruisingen genen kunnen worden overgedragen. Dit betekent dat reproductieve isolatie niet altijd nodig is om een soort te definiëren en dat allopatry (fysieke scheiding) misschien niet nodig is om tot soortvorming te komen.
V: Wat heeft fysieke scheiding te maken met de vorming van nieuwe soorten?
A: Aangenomen wordt dat fysieke scheiding tussen ooit naast elkaar bestaande soorten een belangrijke factor is bij de vorming van nieuwe soorten, zoals blijkt uit vele voorbeelden die in de wetenschappelijke literatuur worden besproken.
Gerelateerde artikelen
Auteur
AlegsaOnline.com Soortvorming Leandro Alegsa
URL: https://nl.alegsaonline.com/art/92570
Bronnen
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833